- Database
- Open access
- Published:
Avian Immunome DB: an example of a user-friendly interface for extracting genetic information
BMC Bioinformatics volume 21, Article number: 502 (2020)
Abstract
Background
Genomic and genetic studies often require a target list of genes before conducting any hypothesis testing or experimental verification. With the ever-growing number of sequenced genomes and a variety of different annotation strategies, comes the potential for ambiguous gene symbols, making it cumbersome to capture the “correct” set of genes. In this article, we present and describe the Avian Immunome DB (Avimm) for easy gene property extraction as exemplified by avian immune genes. The avian immune system is characterised by a cascade of complex biological processes underlaid by more than 1000 different genes. It is a vital trait to study particularly in birds considering that they are a significant driver in spreading zoonotic diseases. With the completion of phase II of the B10K (“Bird 10,000 Genomes”) consortium’s whole-genome sequencing effort, we have included 363 annotated bird genomes in addition to other publicly available bird genome data which serve as a valuable foundation for Avimm.
Construction and content
A relational database with avian immune gene evidence from Gene Ontology, Ensembl, UniProt and the B10K consortium has been designed and set up. The foundation stone or the “seed” for the initial set of avian immune genes is based on the well-studied model organism chicken (Gallus gallus). Gene annotations, different transcript isoforms, nucleotide sequences and protein information, including amino acid sequences, are included. Ambiguous gene names (symbols) are resolved within the database and linked to their canonical gene symbol. Avimm is supplemented by a command-line interface and a web front-end to query the database.
Utility and discussion
The internal mapping of unique gene symbol identifiers to canonical gene symbols allows for an ambiguous gene property search. The database is organised within core and feature tables, which makes it straightforward to extend for future purposes. The database design is ready to be applied to other taxa or biological processes. Currently, the database contains 1170 distinct avian immune genes with canonical gene symbols and 612 synonyms across 363 bird species. While the command-line interface readily integrates into bioinformatics pipelines, the intuitive web front-end with download functionality offers sophisticated search functionalities and tracks the origin for each record. Avimm is publicly accessible at https://avimm.ab.mpg.de.
Background
Ever since the advent of commercial next-generation sequencing platforms in the early 2000s with its associated decrease in sequencing costs [1], the number of DNA sequences increased considerably [2]. Generally, these data become publicly accessible in databases provided by projects focussing on different aspects of biological sequence information [3, 4]. Ensembl [5] and NCBI [6] for instance, have a strong focus on genome annotation with the help of RNA transcript information while UniProt has a pronounced emphasis on protein-coding genes and biological function of proteins. UniProt’s records are either based on manually annotated, non-redundant protein sequences (SwissProt) or on high-quality computationally analysed records, which are enriched with automatic annotation (TrEMBL) [7]. Relying on accurate genome annotations and protein descriptions, Gene Ontology (GO) [8, 9] categorises gene products and fits them into a computational model of biological systems. Their assignment deploys a controlled vocabulary, so-called GO terms, to link genes and gene products to biological processes, cellular components, or molecular functions.
However, genome annotation is not standardised, and each service provider uses their own custom-built annotation pipelines. As a consequence, this often leads to ambiguity in gene names during genome annotation with different gene symbols being given to the same gene or the same gene symbol being given to different, but similar genes. Additionally, since the pre-existing wealth of sequencing information relies on model organisms like human and mouse, there is a strong bias in gene symbols towards those chosen for these species. Particularly for model species, this issue has been partially addressed, for example by the Human Genome Organisation (HUGO) Gene Nomenclature Committee (HGNC) [10], the Vertebrate Gene Nomenclature Committee (VGNC) [11], or the Chicken Gene Nomenclature Consortium [12]. However, this neither guarantees that gene names are harmonised among these consortia, nor does it keep researchers from assigning alternative gene symbols in their annotations, especially when working with non-model species. All these constraints during the data acquisition step can be very challenging to overcome while preparing a gene-related study where choosing the “correct” set of genes is a fundamental and repetitive task for each gene [13]. In other words: “Now that next-generation sequencing (NGS) is commonplace in many laboratories and that efficient bioinformatics toolkits have been developed, knowledge extraction is the bottleneck of genomics” [14].
Gene Ontology’s web-based tool AmiGO 2 [15] offers a convenient entry point for searching and browsing genes and gene products. These are hierarchically organised following the top three categories “biological process”, “cellular component” and “molecular function”. For instance, researchers addressing vision can expand to “biological process/behaviour/visual behaviour” and select genes in this category with the option to further narrow down the results to a focal taxon. Olfaction-related genes can be found under “molecular function/binding/odorant binding”, or vocal learning-related genes under “biological process/behaviour/vocalisation behaviour”. The information for each entry includes the GO term, the source of evidence, and a gene symbol. The gene symbol can then be looked up for further information, such as the nucleotide sequence or the amino acid sequence in Ensembl or UniProt, respectively. This information can be used to prepare gene expression experiments, to conduct gene evolution studies, to look into functional variation, to compare genes between different taxa, or to prioritise targets for gene-editing technologies. These repetitive preparatory steps, however, become the more cumbersome, the more genes are considered.
The immune system in vertebrates is an example of a highly complex biological process [16, 17] represented by more than 25,000 gene products, according to the category “immune system process” in Gene Ontology (filtered for Vertebrata, GO release 2020-06-01). Within the scope of medical research, many immune-related studies have been conducted in human and mouse.
The avian immune system is quite different to that of mammals and is exhibited by different immune organs, cell types and gene repertoires. A fundamental difference between mammals and birds is the absence of lymph nodes but the presence of diffuse lymphoid tissue in birds [18]. As they lack lymph nodes, chickens are thus also missing the genes for the lymphotoxins and lymphotoxin receptors. The lack of functional eosinophils correlates with the absence of the eotaxin genes and the previously reported observation that interleukin-5 (IL-5) is a pseudogene [19].
Other structural and functional differences between the immune systems of birds and mammals include the architecture of the MHC [20] and different modes of somatic recombination in the generation of antibody diversity. Humans have a set of IgM, IgD, IgG, IgA, and IgE antibodies, whereas chickens have IgA, IgM and IgY (this being the equivalent of mammalian IgG).
Some immune genes are either completely missing in birds or are only present in particular species [21] and immune gene families have different orthologues between species, for example, the numbers and types of cytokines and chemokines [22]. It is therefore important that we have a means of cataloguing and a tool for searching these essential genes in avian species.
Immune system-related research in birds is crucial to better understand, for instance, the spread of zoonotic diseases which is not only important in the context of species conservation but also has a potentially great impact on human health and economy (livestock). Birds are highly mobile with some migrating long distances and are thus a potential reservoir and vector for zoonotic diseases [23,24,25,26,27]. Therefore, eco-immunology [28, 29] has gained momentum over the last decade [21, 30, 31].
Recently, the Bird 10,000 Genomes (B10K) Project [32, 33] completed its second phase where the genome of at least one bird species per family was sequenced [Feng et al., in review]. This resulted in 363 bird genomes sequenced on Illumina platforms. We aggregated the immune genes of these genomes to make them publicly and easily accessible in the Avian Immunome DB (Avimm) at https://avimm.ab.mpg.de [34]. This data was complemented with transcript information and amino acid sequences available in Ensembl and UniProt, respectively.
Within the scope of this project, we define the immunome as the set of genes that fall into the category “biological process/immune system process” according to Gene Ontology. However, we are aware of the difficulty of defining what an immune gene is and what is not [35]. Furthermore, we consider our generalist database to be complementary or an entry point to specialist databases such as the international ImMunoGeneTics information system® (IMGT®, [36]) and acknowledge that we rely on immune genes with associated gene symbols that are available on Gene Ontology, Ensembl, or UniProt.
The only model species in birds where the whole immune system has been studied intensively and for a long time is the chicken (Gallus gallus), mostly for agricultural reasons [37]. The chicken immune system is arguably most comprehensively described with 1488 hits in Gene Ontology (GO release 2020-06-01) as compared to other well-studied bird species (five hits in mallard (Anas platyrhynchos), and three hits each in the zebra finch (Taeniopygia guttata) and collared flycatcher (Ficedula albicollis)). This project aims to compile available information about the avian immunome and at the same time disentangle gene symbol ambiguities by linking genes with different names (symbols) to the same unique gene identifier (gene_id in the database). As a consequence, all alternative gene symbols that are found on Ensembl and UniProt are included. Since there is no general standardisation or harmonisation of gene symbols, this intentionally inclusive approach allows for searching our database using different names for the same gene and thus, facilitates comparative studies. Canonical gene symbols (based on Ensembl) are highlighted, and original identifiers of each record are retained and linked to the source provider.
Currently, the database contains evidence for avian immune genes from Ensembl, UniProt and B10K. Avimm strives to become a one-stop resource for avian immunogenomics to facilitate studies in comparative genomics, studies looking into functional variation and the molecular evolution of immune genes and their pathways on whole-genome level [38], as well as an entry point for immune gene expression experiments in birds [39] (Fig. 1). Avimm can be searched by gene symbols and species through the web interface or query script, and the results (lists, annotations, nucleotide and amino acid sequences) can be readily downloaded in various formats.

Concept of Avimm, the Avian Immunome DB
Furthermore, Avimm is a case study to demonstrate the utility of our database layout and implementation for biological processes other than the immune system, for example, olfaction or vocal learning. At the same time, the database concept can be applied to virtually any taxonomic group or rank. An extensive description of the database design is available on our project’s repository [40], which provides scripts and instructions to facilitate the setup of similar databases.
Construction and content
A detailed reconstruction of Avimm is described in our wiki [41]. In brief, the relational database MariaDB (v10.1.44) was installed on a Linux server (Ubuntu 18.04.4 LTS) and secured according to MariaDB’s documentation. The logical data model (LDM) consists of core tables, feature tables and their relationship to each other. The core tables keep track of essential information like species and taxonomy, the origin of evidence and their release cycle versions, gene symbols and descriptions, original evidence identifiers (accession numbers), and resolution of many-to-many relationships (mapping of identifiers). The feature tables contain transcript information, including isoforms, annotation, and nucleotide and amino acid sequences (Fig. 2).

Logical data model (LDM) in Crow’s Foot Notation [42] of Avimm. Core tables represent the basic structure of the database and link the evidence-specific identifiers to Avimm’s unique identifiers (uid). Feature tables represent evidence-specific features like isoforms, nucleotide sequences, or amino acid sequences of the immune genes
Data acquisition and import
As a starting point, genes based on “biological process/immune system process” were selected in AmiGO 2 and further filtered for the best-studied model bird chicken (Gallus gallus, GO release 2020-06-01). These gene symbols were used as a “seed” for creating Avimm. First, the core tables were loaded, including gene synonyms. Feature tables were then subsequently loaded. Detailed scripts and descriptions are available on Avimm’s GitLab repository [40].
By using the gene symbols obtained from GO, Ensembl (release 100, Apr 2020) was queried by a Python script using the Ensembl public MySQL servers [43]. The script queries Ensembl’s REST interface [44] for available species and their current releases [45]. After that, the script checks which of the species are contained in B10K’s current annotation of 363 bird species. For each of these species, the script connects to the corresponding Ensembl database and downloads all Ensembl stable IDs (Ensids) associated with a gene symbol contained in the “seed”. In this process, also the synonyms are checked (table external_synonym in Ensembl). If a gene symbol of the “seed” is found in the synonyms, the corresponding Ensid is downloaded as well. The Ensids were then loaded into Avimm. In the next step, all gene synonyms for the Ensids loaded into Avimm were downloaded from Ensembl. Another Python script was used which connects to each of the relevant species databases and downloads the synonyms for each Ensid contained in Avimm. These synonyms were then also loaded into Avimm.
UniProt data were obtained from the UniProt website. For this purpose, a list of Ensids contained in Avimm was prepared. With this list, the UniProt web interface [46] was queried, and the resulting data were downloaded filtered by the following columns: “Entry (ID)”, “Entry name”, “Status (reviewed/unreviewed)”, “Protein names”, “Organism”, “Gene names (primary)”, “Gene names (synonym)”, “Length”, “Sequence”, “Your list”. As an initial step, only Entry IDs (UniProtKBs) were loaded into Avimm.
For the import of the B10K data, all gene symbols found in Avimm (including synonyms) were used. For each of these symbols, it was checked whether the gene symbol was found in the B10K genomes’ annotations. If the gene symbol was found, the B10K ID (Unigene; B10K-internal identifier and not to be confused with NCBI’s Unigene) was extracted and imported into Avimm.
To fill the feature tables, all obtained accession numbers from Ensembl, UniProt and B10K were then used to extract transcript information, amino acid sequences and nucleotide sequences, respectively, which were subsequently imported into Avimm (Fig. 3). Data were imported via a stored procedure (function in the database) which performs consistency checks before importing data.

Flow chart, loading of Avimm. Uncoloured boxes show preparatory steps. Yellow boxes describe imports into core tables and green boxes imports into feature tables. The initial set of immune genes in chicken is derived from “biological process/immune system process” in AmiGo 2 filtered for chicken. Gene symbols were extracted from Ensembl’s website since Ensembl has the most current/complete chicken annotation. All steps are explained in detail on the project’s wiki page [41]
Synonyms of gene symbols were handled in the following way: The primary gene symbol used in Ensembl was stored as a canonical name in Avimm and identified by a unique gene identifier (gene_id). The synonyms found in Ensembl were also imported into Avimm. Each gene symbol (primary or synonym) received an additional unique gene symbol identifier (uid). This uid was used to identify the gene symbol when storing and extracting data. Synonyms were linked to their canonical name via gene_id. UniProt was queried using the Ensembl stable IDs (Ensids) stored in Avimm. Additional gene symbols found by this process were also stored in Avimm as synonyms with a uid and linked to the canonical name via gene_id. As such, uid uniquely identifies a gene symbol while gene_id identifies a gene. The link between uid and gene_id allows for finding all data related to a gene. For a user it is sufficient to know one of the gene symbols corresponding to a gene (either the canonical name or a synonym); the link between uid and gene_id allows to identify the canonical gene name.
Gene properties in the database can be accessed in two ways
A Python script qimm.py (QueryIMMunome) was written as a command-line interface (CLI) to query data and output nucleotide and amino acid sequences in fasta format [40]. This script can be readily incorporated in bioinformatics pipelines. As a graphical user interface, a web front-end was developed, which allows for easy access to the data in Avimm [34], independent of specialist bioinformatics skills.
Django [47], a widely used web framework built on Python that is known to facilitate the development of web applications, was used for the development of Avimm’s web interface. With both the CLI and the web interface written in Python, it was effortless to reuse the program logic written for the CLI in the web interface. The web interface has two primary purposes: collecting the parameters needed by qimm.py in a user-friendly way and presenting the query results. The parameters collected by the web interface are fed into qimm.py, which queries the database and returns the results, which are then displayed by the web front-end.
The web front-end consists of the main page with a general description of the project and four function pages (Fig. 4), each one corresponding to one of the main modules/functions provided by qimm.py. These function pages provide user-friendly methods to enter query parameters.

Avimm landing page with links to the four function pages (upper box) and description for each function (lower box)
For example, gene symbols are presented in two lists: one containing all available gene symbols, the other the selected gene symbols. Marked symbols can be moved from one list to the other. Besides this, gene symbols can be entered into the text field on the bottom, for example, by copy and paste. Species can be selected by a set of lists representing the taxonomic ranks starting at the class level. Once a selection in an upper level is made, the lists in the lower levels only contain the values compatible with the selection in the upper level. Which data to be displayed can be selected via checkboxes. For other options, for example, downloading data, checkboxes are also provided (Fig. 5).

Excerpt of evidence function page. Gene selection either by “mark-and-arrow” or free text (top left box). Species selection based on taxonomic ranks (centre box). Checkboxes to filter columns (right box) and download data option (bottom left box). Each section has an information button (circled “i”) to provide further details
The web front-end has the additional benefit that hyperlinks can be used. As a consequence, the result page of the evidence function provides direct links to the function pages of Ensembl, UniProt and B10K with the same selection of gene symbols and species that have been used on the evidence function page. Accession numbers in the results of Ensembl and UniProt queries are linked to the corresponding entries within Ensembl and UniProt, respectively. The B10K function page provides links to NCBI BLAST [48] for each result record. The Ensembl function page provides links for each record to the corresponding gene in Ensembl and NCBI. The UniProt function page provides links to the corresponding protein in UniProt and NCBI’s protein blast (BLASTP).
Utility and discussion
The current size of Avimm is 7.3 GB, which encompasses 363 bird species with an average number of 709 immune genes per species [Table 1]. Within the 1782 immune gene records, 363 genes with more than one distinct gene symbol were identified. The gene IL8 (Interleukin-8), for example, is pointing to nine synonymous gene symbols (CEF4, CXCL13, CXCL13L2, CXCL8, EMF1, gIL-8GN, IL8L1, interleukin-8 and k60).
Examples from the CLI
A quick overview of what evidence is available in the database is given below, using the gene IFNL3A (Interferon lambda-3 A) across all species and can be accomplished on the command line:

Explanation of arguments:
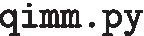 :
:-
the script’s name
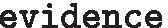 :
:-
query the evidence table
 :
:-
all orders (SQL wild card % for all entries)
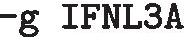 :
:-
the gene of interest
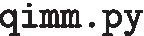 :
: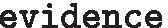 :
: :
: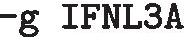 :
:This query will return a species list for which there is evidence for this gene and the type of evidence (B10K, Ensembl or UniProt). Names of alternative gene symbols will be printed at the bottom (in this example, IFNL, IFNL3, IL-28B, IL28 and IL28B are synonyms for IFNL3A). Note that the same results will be returned when querying, for instance, with “-g IL28” since it maps to the same gene in Avimm.
The Ensembl tables need to be queried to retrieve transcript information:

This query will return the Ensembl accession numbers for the genes and transcripts which can then be pasted in Ensembl’s website [49] to get further information.
Amino acid sequences for the gene product are queried through UniProt:

This query will return a fasta-formatted output of the amino acid sequences and adding the argument “-w” to the query will generate fasta output files which can then be used as input for an alignment software for further downstream analyses of these sequences.
Nucleotide sequences of the mRNA can be queried using the B10K module:

The output is again fasta-formatted and adding “-w” will generate fasta files which can be used for downstream analyses of these nucleotide sequences of IFNL3A. A more detailed list of examples, including generated output, can be found on Avimm’s GitLab repository [40].
For instance, a simple gene presence-absence analysis of the gene MASP2 in Avimm,

shows evidence for almost all Anseriformes (5/7) and all Galliformes (11/11), but only for two Passeriformes (2/173) in the database. In humans, MASP2, mannan-binding lectin-associated serine protease-2, is involved in the activation of the lectin pathway of the complement system. MASP2 deficiency was associated with increased susceptibility to infections or autoimmune diseases [50]. The order-biased evidence immediately raises two questions: Is MASP2 mostly absent in Passeriformes, or is it a sequencing (or annotation) artefact? If the former case was true, why is it present in most Galloanserae?
Examples of web front-end
Evidence for the gene IFNL3A (Interferon lambda-3 A) across all species in Avimm can also be found via web front-end [51]. On top of the result list, there are direct links to B10K, Ensembl, and UniProt result pages based on the same search criteria. On the bottom of the page, alternative gene symbols are listed (Fig. 6).

The gene evidence result list shows one record for each evidence of the gene IFNL3A in the selected species. The columns can be sorted (ascending or descending) by clicking on the corresponding column header. On top of the list are links to B10K, Ensembl, and UniProt result pages based on the same search criteria (upper box). If there are alternative gene symbols for the query gene in Avimm, then these are listed on the bottom of the page (lower box)
The B10K link leads to the B10K results page with annotation and further options (filters, data download) including a link to NCBI’s nucleotide BLAST page for each record (Fig. 7a). The Ensembl link leads to Ensembl transcript information, including links to Ensembl’s gene and transcript IDs, and a link to NCBI’s gene search page for each record (Fig. 7b). The UniProt link leads to UniProt protein information and further options (filters, data download) including a link to UniProt’s ID, and a link to NCBI’s protein BLAST for each record (Fig. 7c).

Excerpts of the result pages for IFNL3A in B10K (a), Ensembl (b), and UniProt (c). Each result page offers additional filters and download options. The original identifiers of the source of evidence are retained and linked to the original entries on Ensembl (b, left box) or UniProt (c, left box). Additionally, links to NCBI nucleotide sequence BLAST (a, box), NCBI gene information (b, right box), and NCBI amino acid sequence BLAST (c, right box) are provided for each record
Absence-presence analysis of the gene MASP2 across all 363 species in Avimm shows evidence in almost all Galloanserae (16/18) whereas there is evidence in only two (out of 173) Passeriformes (Fig. 8).

Excerpt of the result page for MASP2 evidence across all 363 species in Avimm. There is evidence in all Galliformes (11/11) and almost all Anseriformes (5/7) but only in two Passeriformes (out of 173 in Avimm)
Scope and context
Avimm is a representation of the currently available information regarding avian immune genes and can potentially economise repetitive, preparatory steps for studies in comparative genomics, gene expression, functional variation and molecular evolution of immune genes. This is ensured by a pre-selection of immune genes based on the well-annotated chicken genome as well as an automated resolution, incorporation and representation of ambiguous gene symbols. A quick presence-absence analysis of immune genes for available species in Avimm will become even more useful with the next data release of the B10K consortium (phase III; at least one representative bird genome per genus covering 2,500 genera and subgenera) [32]. The sevenfold increase of genomes and annotations will also mitigate the current limitation of containing only one or a few bird species representatives per bird family. For each entry, original identifiers (accession numbers of Ensembl, UniProt and B10K) are retained which facilitates the gathering of additional gene information from these providers. The accompanying query script qimm.py can be readily incorporated into bioinformatics pipelines to analyse nucleotide and amino acid sequences in fasta format, for instance, for alignments and SNP identification. The database is already prepared to incorporate immune genes of other vertebrate animal classes to further extend the capability for comparative genomics. For this, a stored procedure takes care of necessary consistency checks before importing new data, which will also be an advantage for the incorporation of immune genes of the forthcoming hundreds of platinum-quality genomes from the Vertebrate Genomes Project [52]. These genomes of birds and other vertebrate classes will be of much higher sequencing quality and read contiguity, which will presumably uncover previously undetected immune genes [53, 54]. Connected to this, particularly in the current version, one should keep in mind that the absence of a particular gene in a species does not necessarily reflect biological reality but might be the consequence of the shortcomings of short-read sequencing [54].
Our generalist database is designed in a way to harbour the common denominator of a focal trait and taxonomic group, mainly a broad range of non-model organisms. It relies on gene symbols found in established projects like Ensembl or UniProt. Specialist databases include in-depth expertise on focal genes and gene products for a given species. For example, IMGT [36] has a strong focus mainly on immunoglobulins and T cell receptors in vertebrates. They also assign gene symbols which then have to be approved by HGNC [10] and VGNC [11] before they are eventually incorporated to other generalist databases. Therefore, we recommend to also consult specialist databases after a pre-selection of focal genes in Avimm. Eventually, genes and gene products with assigned gene symbols will be included within Ensembl and UniProt, and will thus also be included in Avimm.
With the release cycles of Ensembl (every three months) and UniProt (every month) it is challenging to keep up-to-date with the most current annotations; all the more, considering that the number of annotated highly contiguous whole-genomes is ever-growing (five bird genomes in Ensembl release 95 but already 40 in release 100) and the automated annotation pipelines of these service providers rapidly increase the number of correct immune gene annotations. Additional Python scripts are provided on the project’s GitLab repository to facilitate the incorporation of new evidence into Avimm.
An essential feature of the relational database design is the internal mapping of genes and identifiers to unique identifiers through uids and gene_ids. This allows the resolution of many-to-many relationships and, in consequence, is the foundation for allowing queries based on any well-known gene symbol for a single gene.
Conclusions
Currently, Avimm proves to be a valuable resource with which to extract avian immune gene information, mainly from the B10K phase II annotation. With phase III, a total of circa 2500 genomes spanning all bird genera are expected [32]. Although only limited transcript information and amino acid sequences are currently available, Ensembl and UniProt continually annotate genomes that are uploaded to their repositories. These annotations can then be easily added to Avimm. Additionally, VGP has completed or nearly completed 38 of 95 proposed bird genomes of the highest sequencing and assembly quality currently available. These genomes will be annotated, and extracted immune gene information can also be added to the database. It is currently advisable to re-create affected tables in Avimm which is facilitated by Python scripts on the project’s website to reflect the release cycles of Ensembl and UniProt.
The organisation of the data in a relational database and the separation of core and feature tables makes it easy to add additional sources and properties of avian immune genes. For the same reasons, it is also possible to apply the database design to other biological processes such as olfaction, vision or vocal learning. Specialist communities in comparative and functional genomics in their respective disciplines face similar or even identical challenges with genetic information scattered across research consortia and databases, with quite a few gene symbols showing ambiguity. Furthermore, the database design is not limited to birds but can be used to represent genomic data for other taxa with little effort. It is possible to then link databases for different taxa or properties. We provide information on how to use our database model for other taxa or other sets of genes in our wiki. Finally, through the intuitive web interface, data acquisition from international consortia and databases becomes less cumbersome for non-bioinformaticians, who otherwise would need to consult multiple data sources with frequently ambiguous gene symbols.
Availability of data and materials
Change history
30 September 2021
A Correction to this paper has been published: https://0-doi-org.brum.beds.ac.uk/10.1186/s12859-021-04388-x
Abbreviations
- Avimm :
-
Avian Immunome DB
- B10K:
-
Bird 10,000
- BLAST:
-
Basic local alignment search tool
- CDS:
-
Coding sequence
- CLI:
-
Command-line interface
- DB:
-
Database
- Ensid:
-
Ensembl stable ID (accession number)
- GB:
-
Gigabyte
- GO:
-
Gene ontology
- LDM:
-
Logical data model
- MHC:
-
Major histocompatibility complex
- mRNA:
-
Messenger RNA
- NCBI:
-
National Center for Biotechnology Information
- NGS:
-
Next generation sequencing
- REST:
-
Representational state transfer
- RNA:
-
Ribonucleic acid
- SNP:
-
Single-nucleotide polymorphism
- SQL:
-
Structured query language
- uid:
-
Unique (gene symbol) identifier
- Unigene:
-
B10K accession number (not to be confused with Unigene Laboratories)
- UniProt:
-
Universal Protein database
- UniProtKB:
-
UniProt Knowledgebase (accession number)
- VGP:
-
Vertebrate Genomes Project
References
Shendure J, Balasubramanian S, Church GM, Gilbert W, Rogers J, Schloss JA, Waterston RH. DNA sequencing at 40: past, present and future. Nature. 2017;550(7676):345–53. https://0-doi-org.brum.beds.ac.uk/10.1038/nature24286.
GenBank and WGS Statistics. https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genbank/statistics/. Accessed 13 May 2020
Brusic V, Zeleznikow J, Petrovsky N. Molecular immunology databases and data repositories. J Immunol Methods. 2000;238(1):17–28. https://0-doi-org.brum.beds.ac.uk/10.1016/S0022-1759(00)00159-9.
Selzer PM, Marhöfer RJ, Koch O. Biological databases. In: Selzer PM, Marhöfer RJ, Koch O, editors. Applied bioinformatics: an Introduction. Cham: Springer; 2018. p. 13–34. https://0-doi-org.brum.beds.ac.uk/10.1007/978-3-319-68301-0_2.
Cunningham F, Achuthan P, Akanni W, Allen J, Amode MR, Armean IM, Bennett R, Bhai J, Billis K, Boddu S, Cummins C, Davidson C, Dodiya KJ, Gall A, Girón CG, Gil L, Grego T, Haggerty L, Haskell E, Hourlier T, Izuogu OG, Janacek SH, Juettemann T, Kay M, Laird MR, Lavidas I, Liu Z, Loveland JE, Marugán JC, Maurel T, McMahon AC, Moore B, Morales J, Mudge JM, Nuhn M, Ogeh D, Parker A, Parton A, Patricio M, Abdul Salam AI, Schmitt BM, Schuilenburg H, Sheppard D, Sparrow H, Stapleton E, Szuba M, Taylor K, Threadgold G, Thormann A, Vullo A, Walts B, Winterbottom A, Zadissa A, Chakiachvili M, Frankish A, Hunt SE, Kostadima M, Langridge N, Martin FJ, Muffato M, Perry E, Ruffier M, Staines DM, Trevanion SJ, Aken BL, Yates AD, Zerbino DR, Flicek P. Ensembl. Nucleic Acids Res. 2019;47(D1):745–51. https://0-doi-org.brum.beds.ac.uk/10.1093/nar/gky1113.
NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018;46(D1):8–13. https://0-doi-org.brum.beds.ac.uk/10.1093/nar/gkx1095.
UniProt: a worldwide hub of protein knowledge. Nucleic Acids Res. 2019;47(D1):506–15. https://0-doi-org.brum.beds.ac.uk/10.1093/nar/gky1049
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G. Gene ontology: tool for the unification of biology. Nat Genet. 2000;25(1):25–9. https://0-doi-org.brum.beds.ac.uk/10.1038/75556.
Resource The Gene Ontology. 20 years and still GOing strong. Nucleic Acids Res. 2019;47(D1):330–8. https://0-doi-org.brum.beds.ac.uk/10.1093/nar/gky1055.
Home | HUGO Gene Nomenclature Committee. https://www.genenames.org/. Accessed 13 May 2020
Home | Vertebrate Gene Nomenclature Committee. https://vertebrate.genenames.org/. Accessed 13 May 2020
Chicken Gene Nomenclature Consortium. http://birdgenenames.org/cgnc/. Accessed 13 May 2020
Sboner A, Mu XJ, Greenbaum D, Auerbach RK, Gerstein MB. The real cost of sequencing: higher than you think!. Genome Biol. 2011;12(8):125. https://0-doi-org.brum.beds.ac.uk/10.1186/gb-2011-12-8-125.
Zerbino DR, Achuthan P, Akanni W, Amode MR, Barrell D, Bhai J, Billis K, Cummins C, Gall A, Girón CG, Gil L, Gordon L, Haggerty L, Haskell E, Hourlier T, Izuogu OG, Janacek SH, Juettemann T, To JK, Laird MR, Lavidas I, Liu Z, Loveland JE, Maurel T, McLaren W, Moore B, Mudge J, Murphy DN, Newman V, Nuhn M, Ogeh D, Ong CK, Parker A, Patricio M, Riat HS, Schuilenburg H, Sheppard D, Sparrow H, Taylor K, Thormann A, Vullo A, Walts B, Zadissa A, Frankish A, Hunt SE, Kostadima M, Langridge N, Martin FJ, Muffato M, Perry E, Ruffier M, Staines DM, Trevanion SJ, Aken BL, Cunningham F, Yates A, Flicek P. Ensembl 2018. Nucleic Acids Res. 2018;46(D1):754–61. https://0-doi-org.brum.beds.ac.uk/10.1093/nar/gkx1098.
Carbon S, Ireland A, Mungall CJ, Shu S, Marshall B, Lewis S. AmiGO: online access to ontology and annotation data. Bioinformatics. 2009;25(2):288–9. https://0-doi-org.brum.beds.ac.uk/10.1093/bioinformatics/btn615.
Holt RA. Immunogenomics: a foundation for intelligent immune design. Genome Med. 2015;7(1):116. https://0-doi-org.brum.beds.ac.uk/10.1186/s13073-015-0247-y.
Buchmann K. Evolution of innate immunity: clues from invertebrates via fish to mammals. Front Immunol. 2014;. https://0-doi-org.brum.beds.ac.uk/10.3389/fimmu.2014.00459.
Nochi T, Jansen CA, Toyomizu M, Eden WV. The well-developed mucosal immune systems of birds and mammals allow for similar approaches of mucosal vaccination in both types of animals. Front Nutr. 2018;. https://0-doi-org.brum.beds.ac.uk/10.3389/fnut.2018.00060.
Avery S, Rothwell L, Degen WDJ, Schijns VEJC, Young J, Kaufman J, Kaiser P. Characterization of the first nonmammalian T2 cytokine gene cluster: the cluster contains functional single-copy genes for IL-3, IL-4, IL-13, and GM-CSF, a gene for IL-5 that appears to be a pseudogene, and a gene encoding another cytokinelike transcript, KK34. J Interferon Cytokine Res. 2004;24(10):600–10. https://0-doi-org.brum.beds.ac.uk/10.1089/jir.2004.24.600.
Kaufman J, Milne S, Göbel TWF, Walker BA, Jacob JP, Auffray C, Zoorob R, Beck S. The chicken B locus is a minimal essential major histocompatibility complex. Nature. 1999;401(6756):923–5. https://0-doi-org.brum.beds.ac.uk/10.1038/44856.
Magor KE, Miranzo Navarro D, Barber MRW, Petkau K, Fleming-Canepa X, Blyth GAD, Blaine AH. Defense genes missing from the flight division. Dev Comp Immunol. 2013;41(3):377–88. https://0-doi-org.brum.beds.ac.uk/10.1016/j.dci.2013.04.010.
Kaiser P, Poh TY, Rothwell L, Avery S, Balu S, Pathania US, Hughes S, Goodchild M, Morrell S, Watson M, Bumstead N, Kaufman J, Young JR. A genomic analysis of chicken cytokines and chemokines. J Interferon Cytokine Res. 2005;25(8):467–84. https://0-doi-org.brum.beds.ac.uk/10.1089/jir.2005.25.467.
Kraus RHS, Hooft PV, Megens H-J, Tsvey A, Fokin SY, Ydenberg RC, Prins HHT. Global lack of flyway structure in a cosmopolitan bird revealed by a genome wide survey of single nucleotide polymorphisms. Mol Ecol. 2013;22(1):41–55. https://0-doi-org.brum.beds.ac.uk/10.1111/mec.12098.
Jax E. Immunology going wild: genetic variation and immunocompetence in the mallard (Anas platyrhynchos). 2019. https://kops.uni-konstanz.de/handle/123456789/46072. Accessed 5 Oct 2020.
Evseev D, Magor KE. Innate immune responses to avian influenza viruses in ducks and chickens. Vet Sci. 2019;6(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci6010005.
Huang Y, Li Y, Burt DW, Chen H, Zhang Y, Qian W, Kim H, Gan S, Zhao Y, Li J, Yi K, Feng H, Zhu P, Li B, Liu Q, Fairley S, Magor KE, Du Z, Hu X, Goodman L, Tafer H, Vignal A, Lee T, Kim K-W, Sheng Z, An Y, Searle S, Herrero J, Groenen MAM, Crooijmans RPMA, Faraut T, Cai Q, Webster RG, Aldridge JR, Warren WC, Bartschat S, Kehr S, Marz M, Stadler PF, Smith J, Kraus RHS, Zhao Y, Ren L, Fei J, Morisson M, Kaiser P, Griffin DK, Rao M, Pitel F, Wang J, Li N. The duck genome and transcriptome provide insight into an avian influenza virus reservoir species. Nat Genet. 2013;45(7):776–83. https://doi.org/10.1038/ng.2657.
Morris KM, Hindle MM, Boitard S, Burt DW, Danner AF, Eory L, Forrest HL, Gourichon D, Gros J, Hillier L, Jaffredo T, Khoury H, Lansford R, Leterrier C, Loudon A, Mason AS, Meddle SL, Minvielle F, Minx P, Pitel F, Seiler JP, Shimmura T, Tomlinson C, Vignal A, Webster RG, Yoshimura T, Warren WC, Smith J. The quail as an avian model system: its genome provides insights into social behaviour, seasonal biology and infectious disease response. bioRxiv 575332 (2019). https://0-doi-org.brum.beds.ac.uk/10.1101/575332.
Viney ME, Riley EM. From immunology to eco-immunology: more than a new name. In: Malagoli D, Ottaviani E, editors. Eco-immunology: evolutive aspects and future perspectives. Dordrecht: Springer; 2014. p. 1–19. https://0-doi-org.brum.beds.ac.uk/10.1007/978-94-017-8712-3_1.
Demas G, Nelson R. Ecoimmunology. Oxford: Oxford University Press; 2012.
Martin LB, Hawley DM, Ardia DR. An introduction to ecological immunology. Funct Ecol. 2011;25(1):1–4. https://0-doi-org.brum.beds.ac.uk/10.1111/j.1365-2435.2010.01820.x.
Pedersen AB, Babayan SA. Wild immunology. Mol Ecol. 2011;20(5):872–80. https://0-doi-org.brum.beds.ac.uk/10.1111/j.1365-294X.2010.04938.x.
Zhang G. Genomics: bird sequencing project takes off. Nature. 2015;522(7554):34. https://0-doi-org.brum.beds.ac.uk/10.1038/522034d.
Home | B10K Database. https://b10k.genomics.cn/index.html. Accessed 8 July 2020.
Avian Immunome DB. https://avimm.ab.mpg.de. Accessed 14 July 2020.
Heng TSP, Painter MW, Elpek K, Lukacs-Kornek V, Mauermann N, Turley SJ, Koller D, Kim FS, Wagers AJ, Asinovski N, Davis S, Fassett M, Feuerer M, Gray DHD, Haxhinasto S, Hill JA, Hyatt G, Laplace C, Leatherbee K, Mathis D, Benoist C, Jianu R, Laidlaw DH, Best JA, Knell J, Goldrath AW, Jarjoura J, Sun JC, Zhu Y, Lanier LL, Ergun A, Li Z, Collins JJ, Shinton SA, Hardy RR, Friedline R, Sylvia K, Kang J. The Immunological Genome Project: networks of gene expression in immune cells. Nat Immunol. 2008;9(10):1091–4. https://0-doi-org.brum.beds.ac.uk/10.1038/ni1008-1091.
Giudicelli V, Duroux P, Ginestoux C, Folch G, Jabado-Michaloud J, Chaume D, Lefranc M-P. IMGT/LIGM-DB, the IMGT®comprehensive database of immunoglobulin and T cell receptor nucleotide sequences. Nucleic Acids Res. 2006;34(suppl–1):781–4. https://0-doi-org.brum.beds.ac.uk/10.1093/nar/gkj088.
Smith J, Speed D, Law AS, Glass EJ, Burt DW. In-silico identification of chicken immune-related genes. Immunogenetics. 2004;56(2):122–33. https://0-doi-org.brum.beds.ac.uk/10.1007/s00251-004-0669-y.
Kraus RHS, Wink M. Avian genomics: fledging into the wild!. J Ornithol. 2015;156(4):851–65. https://0-doi-org.brum.beds.ac.uk/10.1007/s10336-015-1253-y.
Jax E, Wink M, Kraus RHS. Avian transcriptomics: opportunities and challenges. J Ornithol. 2018;159(3):599–629. https://0-doi-org.brum.beds.ac.uk/10.1007/s10336-018-1532-5.
Mueller RC. Avian Immunome DB. Library Catalog: gitlab.com. https://gitlab.com/rcmueller/immunomedb. Accessed 8 July 2020.
Avian Immunome DB Wiki · Wiki · Ralf C. Mueller / Avian Immunome DB. Library Catalog: gitlab.com. https://gitlab.com/rcmueller/immunomedb/-/wikis/Avian-Immunome-DB-Wiki. Accessed 14 July 2020.
Everest GC. Basic data structure models explained with a common example. In: Proceedings fifth Texas conference on computing systems (1976), pp 18–9.
Public MySQL Server. https://www.ensembl.org/info/data/mysql.html. Accessed 8 July 2020.
Yates A, Beal K, Keenan S, McLaren W, Pignatelli M, Ritchie GRS, Ruffier M, Taylor K, Vullo A, Flicek P. The Ensembl REST API: ensembl data for any language. Bioinformatics. 2015;31(1):143–5. https://0-doi-org.brum.beds.ac.uk/10.1093/bioinformatics/btu613.
EnsEMBL::REST. https://rest.ensembl.org/info/species. Accessed 8 July 2020.
Retrieve/ID mapping. https://uniprot.org/uploadlists/. Accessed 8 July 2020.
The Web framework for perfectionists with deadlines | Django. https://www.djangoproject.com/. Accessed 13 May 2020.
BLAST: Basic Local Alignment Search Tool. https://blast.ncbi.nlm.nih.gov/Blast.cgi. Accessed 13 May 2020.
Ensembl genome browser 100. https://www.ensembl.org/index.html. Accessed 8 July 2020.
Sokolowska A, Szala A, St Swierzko A, Kozinska M, Niemiec T, Blachnio M, Augustynowicz-Kopec E, Dziadek J, Cedzynski M. Mannan-binding lectin-associated serine protease-2 (MASP-2) deficiency in two patients with pulmonary tuberculosis and one healthy control. Cell Mol Immunol. 2015;12(1):119–21. https://0-doi-org.brum.beds.ac.uk/10.1038/cmi.2014.19.
Avian Immunome DB - Evidence. https://avimm.ab.mpg.de/evidence/ Accessed 13 July 2020.
Vertebrate Genomes Project. Library Catalog: vertebrategenomesproject.org. https://vertebrategenomesproject.org. Accessed 13 July 2020.
Rhie A, McCarthy SA, Fedrigo O, Damas J, Formenti G, Koren S, Uliano-Silva M, Chow W, Fungtammasan A, Gedman GL, Cantin LJ, Thibaud-Nissen F, Haggerty L, Lee C, Ko BJ, Kim J, Bista I, Smith M, Haase B, Mountcastle J, Winkler S, Paez S, Howard J, Vernes SC, Lama TM, Grutzner F, Warren WC, Balakrishnan C, Burt D, George JM, Biegler M, Iorns D, Digby A, Eason D, Edwards T, Wilkinson M, Turner G, Meyer A, Kautt AF, Franchini P, Detrich HW, Svardal H, Wagner M, Naylor GJP, Pippel M, Malinsky M, Mooney M, Simbirsky M, Hannigan BT, Pesout T, Houck M, Misuraca A, Kingan SB, Hall R, Kronenberg Z, Korlach J, Sović I, Dunn C, Ning Z, Hastie A, Lee J, Selvaraj S, Green RE, Putnam NH, Ghurye J, Garrison E, Sims Y, Collins J, Pelan S, Torrance J, Tracey A, Wood J, Guan D, London SE, Clayton DF, Mello CV, Friedrich SR, Lovell PV, Osipova E, Al-Ajli FO, Secomandi S, Kim H, Theofanopoulou C, Zhou Y, Harris RS, Makova KD, Medvedev P, Hoffman J, Masterson P, Clark K, Martin F, Howe Kevin, Flicek P, Walenz BP, Kwak W, Clawson H, Diekhans M, Nassar L, Paten B, Kraus RHS, Lewin H, Crawford AJ, Gilbert MTP, Zhang G, Venkatesh B, Murphy RW, Koepfli K-P, Shapiro B, Johnson WE, Palma FD, Margues-Bonet T, Teeling EC, Warnow T, Graves JM, Ryder OA, Hausler D, O’Brien SJ, Howe Kerstin, Myers EW, Durbin R, Phillippy AM, Jarvis ED. Towards complete and error-free genome assemblies of all vertebrate species. 2020. bioRxiv 2020.05.22.110833. https://0-doi-org.brum.beds.ac.uk/10.1101/2020.05.22.110833.
Korlach J, Gedman G, King S, Chin J, Howard J, Cantin L, Jarvis ED. De Novo PacBio long-read and phased avian genome assemblies correct and add to genes important in neuroscience research. 2017. bioRxiv 103911. https://0-doi-org.brum.beds.ac.uk/10.1101/103911
BioDATEN – Bioinformatics DATa Environment. http://www.biodaten.info/. Accessed 13 May 2020.
Acknowledgements
The authors are grateful for the early access to 363 sequenced bird genomes and annotations through the B10K consortium, which sparked the idea for this project. We appreciate the time and effort spent by Dr Veronika Laine and Prof. Dr Kathrine Magor for testing an early version of the web front-end. The authors acknowledge support by the local HPC resources through the core facility SCCKN and the Human-Computer Interaction Lab at the University of Konstanz. The computing infrastructure for the final and productive Avimm webserver was provided by “Gesellschaft für Wissenschaftliche Datenverarbeitung mbH Göttingen” (GWDG), Germany.
Funding
Open Access funding enabled and organized by Projekt DEAL. Contribution to developing the web front-end to database connection by BioDATEN [55]. The funders of this study had no influence on the design nor the results or interpretations therein.
Author information
Authors and Affiliations
Contributions
RM: Drafting and discussion of the initial idea for an avian immune gene database, relational database design and implementation, acquisition and parsing of immune gene data from sources, loading of the initial database, programming of parser scripts, stored procedures and CLI, and manuscript writing. NM: Development of web front-end based on Django and Python, improvement and implementation of database design and stored procedures, improvement and automation of data import, manuscript writing of methods and results in sections for the web front-end. JS: Involved in project concept, provision of immune gene resources, testing of DB and editing of the manuscript. LE: Editing of the manuscript. RIK: Editing of the manuscript. RHSK: Discussion of the initial idea and coordination, provision of funding sources. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised: the name has been updated.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Mueller, R.C., Mallig, N., Smith, J. et al. Avian Immunome DB: an example of a user-friendly interface for extracting genetic information. BMC Bioinformatics 21, 502 (2020). https://0-doi-org.brum.beds.ac.uk/10.1186/s12859-020-03764-3
Received:
Accepted:
Published:
DOI: https://0-doi-org.brum.beds.ac.uk/10.1186/s12859-020-03764-3